Introduction
Chronic environmental stress, including social stress, is known to induce inflammation in the brain, which has been observed in the brains of those with psychiatric or psychosocial behaviors due to brain disorders, such as schizophrenia, dementia, encephalitis, autism, and psychopathy [1-16]. In humans, chronically and repetitively abused or maltreated children often develop antisocial behavior, such as aggressive or violent and cruel behaviors. In addition, humans exposed to maltreatment in childhood are at increased risk of conduct disorders, personality disorders, major depression, posttraumatic stress disorder, schizophrenia, and anxiety disorders [17-19]. Thus, a suitable animal model of and experiments on child maltreatment or abuse are necessary for furthering the understanding of the pathogenesis and treatment of related brain disorders [7,17,19-22].
Studies in animals and humans have revealed that early-life stress affects brain development and social behavior in adulthood. In humans, early-life stress (child and adolescent abuse and neglect) can induce brain inflammation and structural or functional brain alterations. Several animal models have been developed to study the effects of early-life social stress on adulthood by subjecting the animals to repeated maternal separation (MS), social isolation (SI), or peripubertal social defeat (SD) protocols [7,17,19-22].
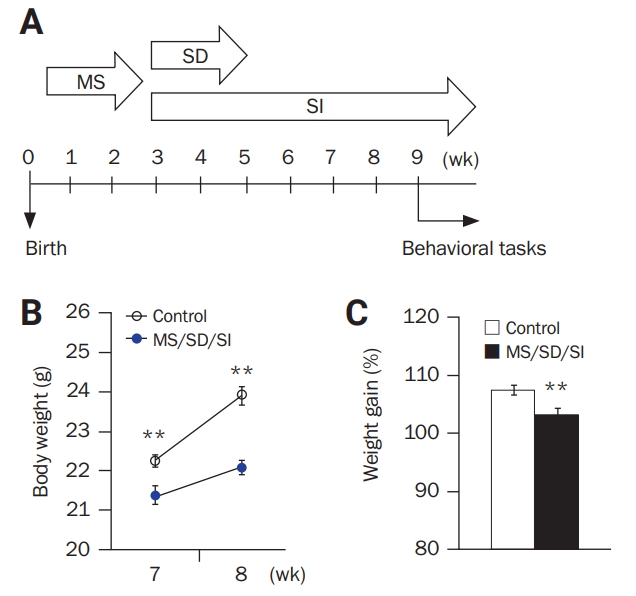
To study the relationship between social stresses in early childhood development (a period of the human lifespan between infancy and adolescence) and social behavior in adulthood, we subjected mice to a sequential process of MS/SD/SI, which is designed to imitate child abuse, and characterized their behavioral phenotypes (Figure 1A).
Methods
Animals
C57BL/6 mice were used in the present study. Animals were maintained with free access to food and water under a 12-hour light/dark cycle. All experiments were approved by the Institutional Animal Care and Use Committee of Seoul National University Hospital (No. 14-0253-S1A1), and all animals were maintained in a facility accredited by Association for Assessment and Accreditation of Laboratory Animal Care (AAALAC) International (No. 001169) in accordance with the Guide for the Care and Use of Laboratory Animals 8th edition (National Research Council 2011). All efforts were made to minimize suffering.
Generation of an MS/SD/SI model
Male offspring of a C57BL/6 female mouse mated with a C57BL/6 male mouse were used in this study. To mimic early-life social stresses, the pups sequentially underwent MS, SD, and SI experiences. The social stress protocols of MS, SD, or SI were applied as described previously [7,17,19-22]. Pregnant females were checked daily for newborns. For each litter, the day of birth was defined as postnatal day (P) 0. The MS procedure was performed from P4 to P18. During this period, litters were separated from their mothers for 3 hours per day. During the period of SD, a second social stress paradigm was introduced to the same mice from P21 to P34. During this period, the mice were singly placed into the cages of singly housed aggressor C57BL/6 mice (territorial residents) for 10 minutes per day for 10 days. Adult 8- to 15-week-old male mice that showed high intermale aggression in a resident-intruder paradigm were selected and used as aggressor mice. As male mice defend their territory against intruding males, the aggressor residents usually initiated a fight. At the start of the SD procedure (P21), the mice were separated from their littermates and housed singly (SI) for the duration of the experiments. The behavioral experiments were started at the age of 9 weeks (usually young adult starts at the aged of 8–9 weeks). The procedure is illustrated in Figure 1A.
Behavioral tasks
Following the sequential experiences of MS, SD, and SI, the MS/SD/SI mice performed behavioral tasks to assess anxiety, locomotor activity, learning/memory, social behavior, aggression, fear, and despair. The behavioral tests were video-recorded and conducted between 13:00 and 18:00 under a light intensity of 80 lux. There were no experimenters in the room during the behavioral tasks, which comprised elevated plus maze, light/dark transition, marble-burying, nest-building, open-field, classical fear-conditioning, observational fear-conditioning, predator fear, resident-intruder, social interaction with a juvenile mouse, sociability, and forced swim (FST) and tail-suspension tests (TSTs). Data were quantified by an experimenter blinded to the conditions. Behavioral experiments were performed as described previously [23-29].
The elevated plus maze was made of plastic and consisted of two white open arms (25 × 8 cm), two black enclosed arms (25 × 8 × 20 cm), and a cross-shaped central platform (8 × 8 × 8 cm). The maze was placed 50 cm above the floor, and mice were individually placed in the center with their heads directed toward one closed arm. The total time spent in each arm or in the center and the total number of entries into each arm were analyzed by video monitoring for 5 minutes. When all four paws crossed from the center into an arm, it was counted as an arm entry.
The light/dark transition test was performed using a plastic light/dark box (30 × 45 × 27 cm) composed of a dark compartment (1/3 of the total area) and a light compartment (2/3 of the total area), with a hole between the two. The light compartment was illuminated at 400 lux. The elapsed time to entry into the light compartment (latency) and the amount of time (duration) spent in each compartment were measured over a 5-minute period by video monitoring.
In contrast, the open-field box was made of white plastic (40 × 40 × 40 cm). Individual mice were placed in the periphery of the field, and the paths of the animals were recorded with a video camera. The total distance traveled for 30 minutes was analyzed using EthoVision XT (Noldus, Wageningen, Netherlands).
For the FST, mice were placed individually in 2,000-mL glass beakers filled with nearly 1,400 mL of water (10 cm from the ground, with a water temperature of 25°C ± 1°C) and were allowed to swim freely for 6 minutes. The duration of immobility, defined as lack of movement in a floating state or performance of only minimal movement required for floating (e.g., small, slow kicking of one paw only) but the absence of active swimming behavior, was measured during the last 4 minutes of the task.
During the TST, mice were hung upside down from a horizontal bar by the tail. Mouse behavior was recorded for 6 minutes, and the time spent immobile was measured.
For classical fear-conditioning, the mice were habituated in a fear-conditioning apparatus chamber for 5 minutes and then subjected to a 28-second acoustic conditioned stimulus followed by a 0.7-mA shock (unconditioned stimulus) applied to the floor grid for 2 seconds (Panlab SLU, Barcelona, Spain). Conditioned stimulus–unconditioned stimulus coupling was carried out three times at 60-second intervals. To assess contextual memory, the animals were placed back into the training context 24 hours after the stressful experiences. The duration of their fear response (freezing behavior) was measured for 4 minutes. To assess the cued memory, the animals were placed in a different context (a novel chamber) for the following 24 hours, and their behavior was monitored for 5 minutes. During the last 3 minutes of this test, the animals were exposed to the conditioned stimulus. Their duration of freezing behavior was measured throughout the 3-minute test.
During the marble-burying test, a single subject mouse was allowed to roam freely in a clean cage (22 × 19 × 17 cm, with a 5-cm sawdust layer) for 10 minutes for habituation. Then, the mouse was removed while 20 glass marbles (15 mm in diameter) were randomly placed in the cage; the subject mouse was reintroduced and allowed to roam freely for 20 minutes. The number of marbles buried (defined by > 50% of the marble being covered by bedding material) was recorded.
For the nest-building test, each mouse was briefly moved to a cage with wood-chip bedding and one Nestlet (Ancare, Bellmore, NY, USA) approximately 1 hour before the dark phase. The following morning, we assessed the nest-building status on a rating scale of 1 to 5 points, as follows: 1, the Nestlet was largely untouched (> 90% intact); 2, the Nestlet was partially torn up (50%–90% remaining intact); 3, the Nestlet was mostly shredded (< 50% remaining intact), but there was no identifiable nest site; 4, there was an identifiable but flat nest (walls higher than mouse height but < 50% of the nest circumference); and 5, there was a well-formed nest with a crater (walls higher than mouse height and ≥ 50% of the nest circumference).
During the sociability task, a three-chambered, white, rectangular plastic box (40 × 25 × 18 cm) was used, into which a wire mesh cage (diameter, 8 cm; height, 10 cm) was placed in each of two side chambers (facing diagonally in the box). Following a 10-minute habituation period, a novel mouse (stranger, age-matched male C57BL/6 mouse) was enclosed in one of the wire cages, and the subject mouse was placed in the middle of the box and allowed to explore for 10 minutes. The time spent in each chamber was measured.
For the social interaction task, a single subject mouse in its home cage was allowed to roam freely for 10 minutes (habituation). A novel juvenile (3–4 weeks old) male C57BL/6 mouse was introduced into the home cage of the resident subject, and both were allowed to roam freely for 5 minutes (test session). The following types of behavior shown by the resident mouse were scored as social interaction: sniffing, grooming, closed chasing, pushing the snout or head under, and crawling over or under the juvenile’s body, as well as mounting. The total time spent performing social interaction behaviors was quantified.
The isolation-induced resident-intruder aggression test was performed by introducing an intruder mouse into the home cage of the resident mouse. Resident mice were housed in isolation for 7 days without a bedding change before testing. Aged-matched, naïve male C57BL/6 mice were used as counterparts. The offensive behavior of the resident (offensive aggressiveness) or intruder mouse (defensive aggressiveness) was measured by the latency to the first attack and the total number of attacks (biting and wrestling) by either for 15 minutes.
For observational fear-conditioning, the apparatus consisted of a dual-channel modular shuttle box consisting of two identical chambers (21 × 17.5 × 25 cm each) and a stainless-steel rod floor (5-mm–diameter rods, spaced 1 cm apart; Med Associates, Albans, VT, USA). A transparent plexiglass partition was placed between the chambers. Briefly, the observer and demonstrator mice (age-matched male C57BL/6 mouse) were habituated in the apparatus chambers for 5 minutes. Then, a 2-second foot shock (1.3 mA) was administered to the demonstrator every 10 seconds for 3 minutes by a computer-controlled shocker (Med Associates). The duration for which the observer mouse displayed freezing behavior (fear response) was measured.
Finally, the predator-avoidance test was designed by modifying the social approach-avoidance test or social target test. The test area was a white acryl plastic open-field box (40 × 40 cm) that contained two triangular wire mesh cages (21 × 21 × 28 cm) located diagonally across each end of the field. One cage (target) contained a social target animal (male rat, 5–10 weeks), and the other cage (no target) was empty. Video-tracking data were used to determine the times spent by the experimental mouse in the “rat zone” and “empty zone” (28 × 6 cm) in front of each cage.
Statistical analysis
All data are presented as mean ± standard error of the mean. Analysis of variance (ANOVA) was used to conduct multiple comparisons of means. The Student t-test was performed to determine statistical differences between the two means. A p-value of < 0.05 was considered to indicate statistical significance.
Results
MS/SD/SI mice showed reduced body weight gain
The MS/SD/SI mice (n = 13) showed lower body weight at 7 and 8 weeks of age (Figure 1B). At 8 weeks, MS/SD/SI mice had gained significantly less weight than the control mice (n = 15), suggesting their slower rate of weight gain (MS/SD/SI, 103.33% ± 1.01% vs control, 107.48% ± 0.85%; p < 0.01, Student t-test) (Figure 1C).
MS/SD/SI mice showed normal anxiety levels and increased locomotor activity
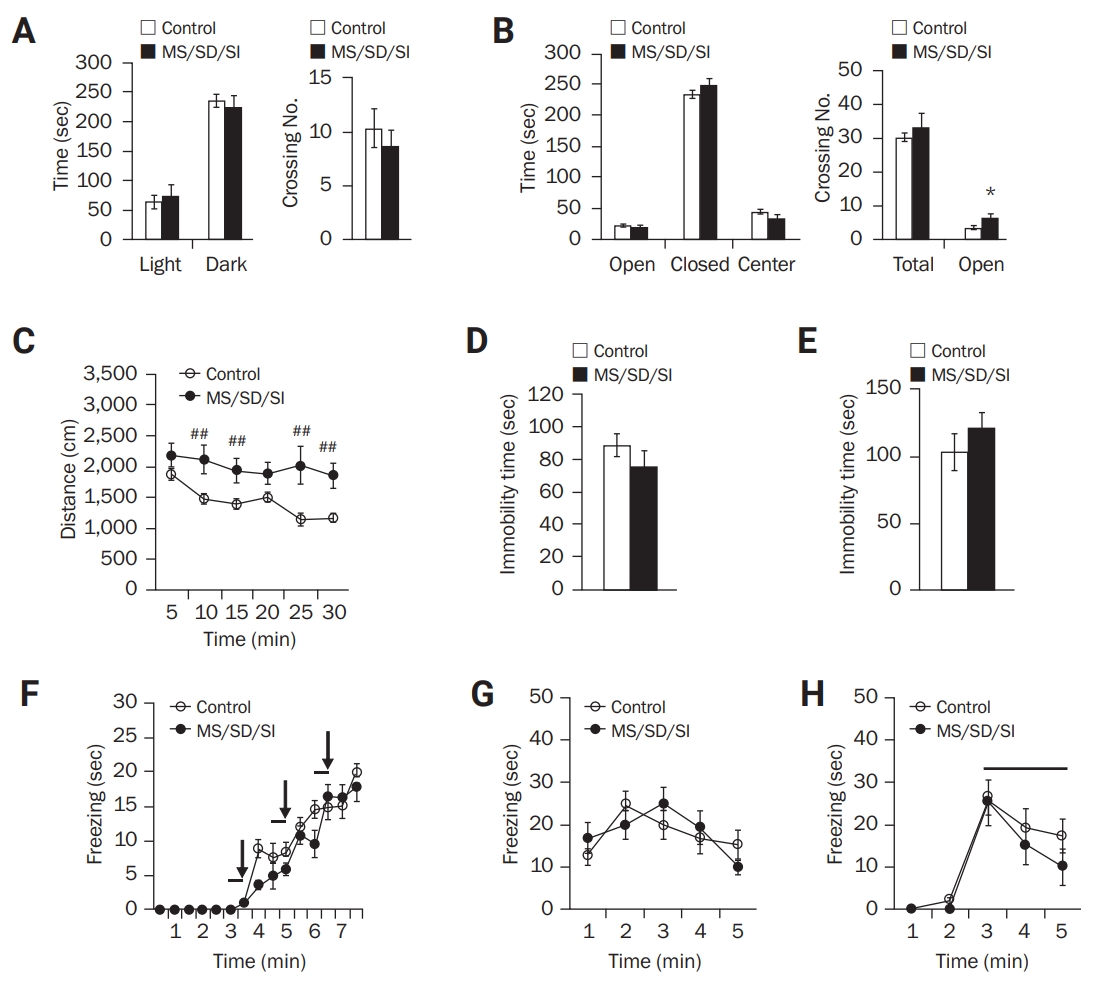
We first subjected the MS/SD/SI mice to the light/dark and elevated plus maze tests to assess anxiety. During the light/dark task (Figure 2A), the MS/SD/SI mice (n = 18) spent a similar amount of time in the dark and light chambers to the control mice (n = 12). In addition, there was no significant difference in time spent in the open and closed arms of the elevated plus maze between the MS/SD/SI (n = 9) and control mice (n = 16) (Figure 2B). However, the MS/SD/SI mice (6.22 ± 1.39 crossings) displayed an increased number of crossings into the open arms compared to the control mice (3.31 ± 0.56 crossings; p < 0.05, Student t-test) (Figure 2B).
To assess locomotor activity, the open-field test was performed, and an increase in distance moved was observed among the MD/SD/SI mice (n = 23) compared to the control mice (n = 14) (F1,35 = 8.70; p < 0.01, two-way ANOVA) (Figure 2C). These results indicate normal anxiety levels but increased locomotor activity among the MS/SD/SI mice compared to the control mice.
MS/SD/SI mice displayed normal behavior in depression- and learning/memory-related tasks
The FST and TST were used to assess behavioral despair, an indicator of depression-like behavior in mice. The durations of immobility of the MS/SD/SI mice during the FST (MS/SD/SI, n = 12; control, n = 15) (Figure 2D) and TST (MS/SD/SI, n = 10; control, n = 12) (Figure 2E) were similar to those of the control mice, indicating a normal level of despair among the MS/SD/SI mice.
For learning and memory tasks, classical fear-conditioning was performed. During the training (Figure 2F), there was no difference in the duration of freezing between the MS/SD/SI (n = 12) and control mice (n = 13), indicating a normal learning ability for fear acquisition of the MS/SD/SI mice. In addition, the MS/SD/SI mice exhibited similar freezing durations in 24-hour contextual (Figure 2G) and cued memory tests (Figure 2H) to those of the control mice, which indicates that the MS/SD/SI mice normally formed memories of fear.
MS/SD/SI mice exhibited altered repetitive/compulsive-like behavior
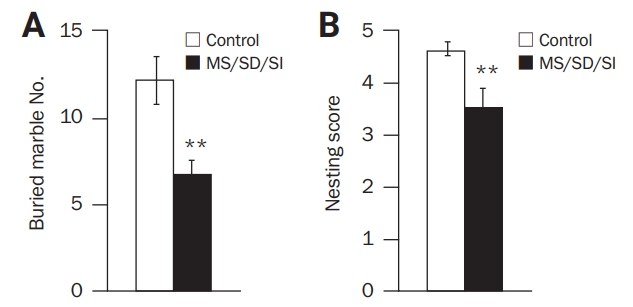
To investigate repetitive/compulsive-like behavior, marble-burying and nest-building performance tests were conducted. During the marble-burying test, the MD/SD/SI mice (n = 9) showed a decrease in the number of buried marbles (6.78 ± 0.78 marbles) compared to the control mice (n = 11, 12.18 ± 1.37; p < 0.01, Student t-test) (Figure 3A). In nest-building behavior, a low score was recorded for the MD/SD/SI mice compared to the control mice (n = 10, 3.50 ± 0.37 points vs n = 13, 4.62 ± 0.13 points, respectively; p < 0.01, Student t-test) (Figure 3B). These results demonstrate that MD/SD/SI mice show lower levels of repetitive/compulsive-like behaviors than control mice.
MS/SD/SI mice exhibited abnormal and unique social behaviors
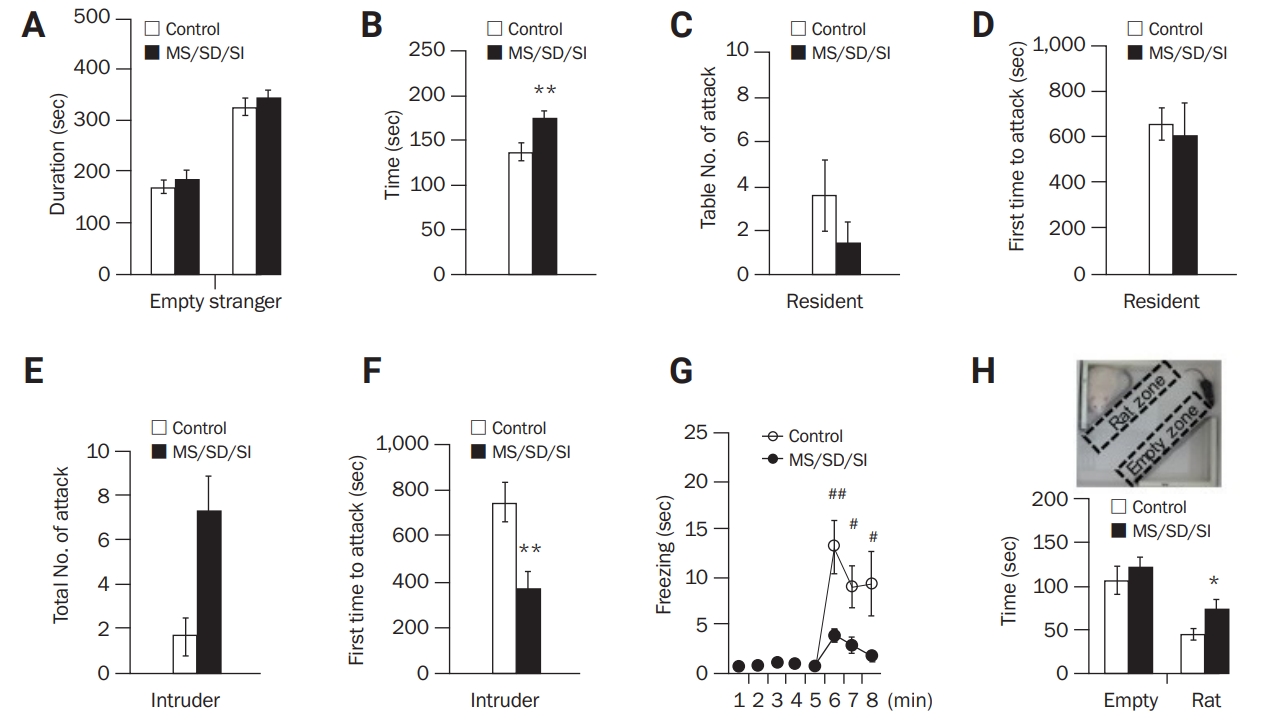
To investigate social behaviors, tasks related to sociability, social interaction, aggression, and social fear were conducted. During the sociability task, the amount of time spent on each side of the chamber containing an empty or stranger cage did not differ between the MD/SD/SI (n = 11) and control mice (n = 16) (Figure 4A). However, in a social interaction task with a juvenile mouse, the MD/SD/SI mice showed significantly higher interaction times than the control mice (n = 16, 173.85 ± 8.20 seconds vs n = 20, 136.31 ± 9.93 seconds; p < 0.01, Student t-test) (Figure 4B). The social interaction behaviors of the MD/SD/SI mice were further investigated using a resident-intruder test related to aggression. When the MD/SD/SI mice were positioned as residents for examination of offensive behavior, they (n = 12) showed a similar number of attacks (Figure 4C) and latency before the first attack (Figure 4D) to those of control mice (n = 18). However, when the MD/SD/SI mice were positioned as intruders to assess defensive aggressiveness, they (n = 16) exhibited a greater number of attacks (MS/SD/SI, 7.25 ± 1.63 attacks vs control, 1.63 ± 0.84 attacks) (Figure 4E) and a shorter latency before the first attack (MS/SD/SI, 370.24 ± 75.80 seconds vs control, 748.75 ± 87.78 seconds; p < 0.01, Student t-test) compared to the control mice (n = 8) (Figure 4F), indicating a higher level of defensive aggressiveness in the MD/SD/SI mice.
For assessment of social fear, empathy-related observational fear-conditioning and innate fear-related predator-avoidance tests were performed. Regarding observational fear-conditioning, the MD/SD/SI mice (n = 12) exhibited a shorter freezing duration than the controls (n = 12) (F1,22 = 4.09; p < 0.05, two-way ANOVA) (Figure 4G), indicating impaired observational fear learning in the MD/SD/SI mice. In the predator-avoidance task, both the MD/SD/SI (n = 22) and control mice (n = 17) spent less time in a rat zone (an area containing a rat) than in an empty zone. However, there was a significant difference in the amount of time spent in the rat zone between the MS/SD/SI and control mice (74.94 ± 10.25 seconds vs 45.63 ± 6.93 seconds, respectively; p < 0.05, Student t-test) (Figure 4H), indicating a dysfunction of predator fear in the MS/SD/SI mice.
Taken together, these results demonstrate that MD/SD/SI mice display unique behavioral features that show impairment of social fear as well as high levels of social interaction and aggression.
Discussion
In this study, we subjected mice to a sequential process of early-life social stress or adverse events of MS, SD, and SI, imitating human child abuse or maltreatment. The MS/SD/SI mice showed normal behaviors in anxiety-, depression-, and learning/memory-related tasks, but they exhibited profound abnormalities in social behaviors.
The MS/SD/SI stress paradigm had a similar result as chronic stress is known to slow weight gain [30]. However, the MS/SD/SI mice showed distinct phenotypes in some behavioral experiments compared to the animal models created by a single-social stress paradigm of either MS, SD, or SI. It has been reported that animal models generated by the social stress paradigm of either MS or SD or SI usually exhibit more depressed and anxious behaviors than control animals [22]. In addition, the single-paradigm models generally had a reduced level of social interaction or sociability, except an adolescent SI model [22,31]. Even mouse models that are known to display aggressive behaviors instead exhibit offensive aggressiveness when subjected to such stresses. Moreover, little is known about the empathy-related behavior of such animals. However, our MS/SD/SI mice showed normal behaviors in anxiety- and depression-related tasks but increased social interaction and defensive aggressiveness. Since they showed normal behavior during the sociality test, it is not likely that their increased level of social interaction is due to a simple altered response to a novelty or curiosity. In the resident-intruder test, the intruders usually show defensive behaviors in response to offensive attacks by the territorial resident. However, MS/SD/SI mice attacked the residents while being intruders, seeming to take the role of dominant in the interaction. The MS/SD/SI mice showed impairment in the expressions of predator fear and empathy-related fear but not those of hippocampus- and amygdala-dependent conditioned fear. These results suggest that our MS/SD/SI stress paradigm affects brain substrates or circuits, such as the frontal cortex, involved in social fear expression and empathetic or aggressive emotion [32-41].
Regarding the hyper-phenotypes in social interaction and offensive aggressiveness and the hypo-phenotypes in predator fear and empathy-related behavior of MS/SD/SI mice, further study of antisocial behavior is needed to characterize the phenotypes in more detail. The observed behavioral traits in the presented model suggest MS/SD/SI mice as a model of personality disorders, such as psychopathy, given the characteristics of antisocial behavior, impaired empathy, and excessive boldness.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print