Chronic social stress during early development is involved in antisocial maltreatment behavior in mice
Article information

Abstract
Purpose
Early-life stress can cause brain inflammation and affect social behavior in adulthood. In humans, maltreated (abused or neglected) children often exhibit antisocial behavior, including violent and sadistic behavior, in adulthood. However, it is unknown whether maltreatment behavior occurs in rodents. Here, we developed an assay system to evaluate conspecific maltreatment behavior in the mouse.
Methods
To assess maltreatment behavior, we devised a two-chamber apparatus separated by a transparent partition, in which one chamber was provided with a nose-poking hole that would trigger foot shocks onto the other. Lidocaine was used to inhibit neural activity in vivo. Brain oscillations were investigated by electroencephalograph. Enzyme-linked immunosorbent assay was used for protein assay. The mouse model was sequentially subjected to maternal separation (MS), social defeat (SD), and social isolation (SI) in that order (MS/SD/SI model).
Results
Inactivation of the anterior cingulate cortex and medial prefrontal cortex increased the level of nose-poking. Maltreatment behavior provoked changes in oxytocin, corticosterone, and brain-derived neurotrophic factor levels. MS/SD/SI mice exhibited more sustained nose-poking behavior during the experiment, resulting in increased foot shocks to the mouse in the opposite chamber. Abnormal brain oscillations were observed in the MS/SD/SI mice.
Conclusion
The MS/SD/SI model and maltreatment-behavior assay may be useful not only to study the relationship between social stress in childhood and antisocial behavior in adulthood, but also for study of etiology, pathology, or treatment for brain disorders, such as psychopathy.
Introduction
Chronic environmental stress is known to induce inflammation in the brain, which is closely related to psychiatric or psychosocial behaviors shown in brain disorders such as schizophrenia, dementia, encephalitis, autism, and psychopathy [1-27]. In humans, repeatedly abused or maltreated children often fail to develop good social relationships and display antisocial behaviors, such as aggressive or violent behaviors, as adults. Furthermore, child maltreatment is associated with increased risks for many disorders including conduct disorder, personality disorder, major depression, posttraumatic stress disorder, schizophrenia, and anxiety disorder [28-30]. Empathy deficit is closely related to antisocial and maltreatment behaviors [31-36], and recent animal studies revealed that emotion contagion or empathy and prosocial behavior occur in rodents [37-40]. However, maltreatment behavior induced by empathy deficit has never been examined in rodents.
Studies in animals and humans have revealed that early-life stress affects brain development and social behavior in adulthood [7,28,30,41-43]. In fact, early-life stress (child and adolescent abuse and neglect) can induce structural or functional alterations in the brain [44-52]. In a previous study, we characterized behavioral phenotypes of a mouse model generated by a sequential process of early-life social stress imitating child abuse or maltreatment composed of maternal separation (MS), social defeat (SD), and social isolation (SI). The MS/SD/SI mice exhibited hyper-phenotypes of social interaction and offensive aggressiveness and hypo-phenotypes of predator fear and empathy-related behavior.
In this study, we developed an assay system to evaluate maltreatment behavior using a perpetrator-victim paradigm. The system consists of a two-chamber apparatus separated by a transparent partition, in which one chamber was provided with a nose-poking hole that triggered foot shocks to a conspecific in the other chamber (Figure 1A). In order to study the relationship between early-life social stresses and antisocial maltreatment behavior, we subjected MS/SD/SI model mice to this assay system and investigated rhythmic brain activities during maltreatment behavior by electrophysiological assessment in vivo.
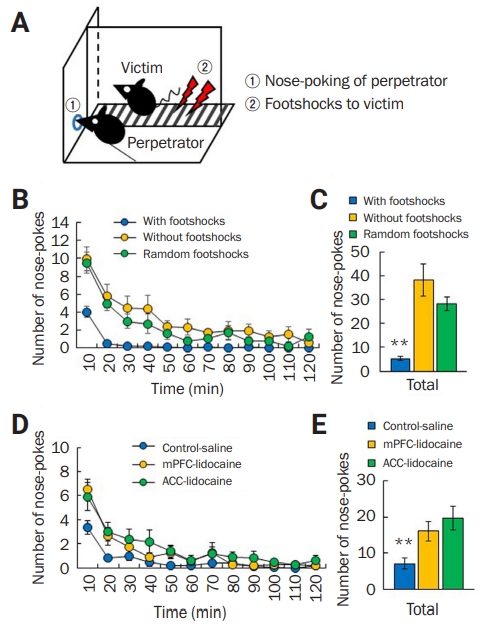
Maltreatment-behavior task
(A) Schematic drawing of the maltreatment-behavior task. (B, C) Number of nose-pokes by normal male C57BL/6 mice (perpetrator) in the presence (with, n = 17) or absence (without, n = 16) of foot shocks or in the condition of random (random, n = 10) foot shocks to victim mice every 10 minutes for 120 minutes. (B) Average number of nose-pokes for a total of 120 minutes. (C) C57BL/6 perpetrator mice showed a low number of nose-pokes in the condition with foot shocks compared to other conditions. **p < 0.01, one-way analysis of variance (ANOVA). (D, E) In the condition with foot shocks, administration of lidocaine into the medial prefrontal cortex (mPFC, n = 11) and anterior cingulate cortex (ACC, n = 11) increased the number of nose-pokes by the perpetrator mice compared to those receiving saline injections (n = 16). **p < 0.01, one-way ANOVA.
Methods
Animals
Animals were maintained with free access to food and water under a 12-hour light/dark cycle. All experiments were approved by the Institutional Animal Care and Use Committee at Seoul National University Hospital (No. 14-0253-S1A1), and all animals were maintained in a facility accredited by Association for Assessment and Accreditation of Laboratory Animal Care International (No. 001169) in accordance with Guide for the Care and Use of Laboratory Animals, 8th edition (National Research Council 2011). All efforts were made to minimize suffering. C57BL/6, BALB/c, and 129S4/Sv mice were used in the present study.
The apparatus and behavioral task for maltreatment behavior
The apparatus for maltreatment behavior was constructed by modifying a shuttle box used for an avoidance test (Med Associates, Fairfax, VT, USA). The modified apparatus consisted of two chambers partitioned by a transparent polymethyl methacrylate divider and a stainless-steel grid rod floor (5-mm diameter rods, spaced 0.7 cm apart): one chamber was designated as the victim compartment (10 × 6 × 21 cm), and the other chamber as the perpetrator compartment (20 × 10 × 21 cm). The grid floor in the victim compartment was exposed, whereas an opaque acrylic plate (20 × 10× 0.3 cm) was placed on the grid floor in the perpetrator compartment in order to prevent electric shocks. The perpetrator compartment contained a nose-poke hole near the transparent divider, comprised of an infrared control, source, and detector. Whenever the beam of the nose-poke was broken by entry of the mouse’s (perpetrator’s) nose from the perpetrator compartment, a 2-second foot shock (1 mA) was delivered to the mouse (victim) in the victim compartment through a grid floor via a computer-controlled animal shocker. We chose 1 mA foot-shock intensity because it resulted in a homogeneous response by victim mice including running, vocalization, and jumping. Thus, perpetrator mice received similar inputs from all victim mice during the maltreatment behaviors. The number of nose-pokes (equal to the number of beam breaks and the number of electric shocks delivered) was recorded in real time, analyzed every 10 minutes, and used for assessing levels of maltreatment behavior. All modules and paradigms were programmed by the MED-SYST-8 interface and software package (Med Associates).
To assess maltreating behavior, mice (perpetrator and victim) were initially individually placed in their apparatus chambers for 10 minutes. During the 10-minute habituation, the nose-poke hole was covered with a panel the same as that used for walls in the chamber. When the panel was removed after the 10-minute habituation, the perpetrator mouse could easily find the nose-poke hole. Each experiment was conducted for 120 minutes. Electric shocks were only delivered to a victim mouse at the moment when the perpetrator mouse put its nose into the hole (triggering foot shocks). To optimize the maltreatment behavior, we performed control experiments and obtained a behavior profile of perpetrator mice: (1) the victim mice were given no foot shocks (without foot shocks) or (2) the victim mice were given random foot shocks regardless of nose-poking (random foot shocks). In the random shock condition, the total number of electrical shocks was the same as the average number of nose-pokes in the foot-shock condition. Unless otherwise stated, we used male C57BL/6 mice in all experiments.
Generation of the maternal separation/social defeat/social isolation model
The MS/SD/SI model was generated as described previously [53]. Male offspring of a C57BL/6 female mouse mated to a C57BL/6 male mouse were used in this study. To mimic early-life social stresses, pups sequentially underwent MS, SD, and SI procedures. Pregnant females were checked for litters daily. If litters were found, the day of birth was defined as postnatal day (P) 0 for that litter. The MS procedure was implemented from P4 to P18. During this period, litters were first separated from their dams and placed in a new cage for 3 hours per day and then were returned to their home cages. The SD, a second social stress paradigm, proceeded for the same mice from P21 to P34. During this period, the mice were singly placed as intruders into the cages of singly housed aggressor C57BL/6 mice (territorial residents) for 10 minutes per day for 10 days between P21 and P34. Adult 8- to 15-week-old male mice that show high level of intermale aggression in a resident-intruder paradigm were selected and used as aggressor mice. As male mice defend their territory against intruding males, the aggressor residents usually initiated a fight. At the beginning of the SD procedure (P21), the mice were separated from their littermates and housed singly (SI) until all experiments were complete. All experiments were started at the age of 9 weeks. The MS/SD model was generated only by MS and SD without SI, and the SI model was generated only by SI, without MS and SD.
Cannula implantation and microinjection
In vivo microinjection and surgery were performed as previously described [39]. For microinjection, a plastic cannula (Virginia Plastics Co., Roanoke, VA, USA) was implanted in the right hemisphere at anteroposterior (AP) +1.5 mm, lateral (L) 0.2 mm, and dorsoventral (DV) 1.25 mm from bregma in the anterior cingulate cortex (ACC) and at AP +2.0 mm, L 0.2 mm, and DV 2.0 mm from bregma in the medial prefrontal cortex (mPFC) using a stereotaxic apparatus (Kopf Instruments, Tujunga, CA, USA). Experiments began 7 days after surgery. Lidocaine (4%, vol/vol, 0.5 μL) or saline (0.9% NaCl, vol/vol, 0.5 μL) was infused into each brain area via an inner cannula (33 gauge) connected to a 25-μL Hamilton syringe. The flow rate (0.1 μL/min) was regulated by a syringe pump (SP100i, WPI). After finishing all experiments, the position of the cannula was histologically verified.
In vivo electrophysiology for electroencephalography
In vivo electroencephalography (EEG) and surgery were performed as previously described [39,54]. To monitor brain oscillations, the mice were subjected to EEG surgery 2 weeks before the maltreatment behavior experiment. For surgery, the animals were anesthetized by intraperitoneal injection of ketamine (90 mg/kg) and xylazine hydrochloride (40 mg/kg). Electrode implantation was performed using a stereotaxic apparatus (Kopf Instruments). EEG recordings were obtained using skull screws (stainless steel, 1.0 mm in diameter), which were positioned −1.94 to 1.70 mm AP and 0.4 to 0.5 mm ML from bregma with grounding over the cerebellum. The electrodes were fixed to the skull with cyanoacrylate adhesive and dental acrylic cement. EEG recordings combined with video monitoring were performed simultaneously during the behavioral task of maltreatment behavior by the perpetrator mouse. A 6-second signal immediately before and after the second nose-poke of a perpetrator mouse was selected and used for analysis. Five EEG frequency-bands—delta (1.5–4 Hz), theta (4–8 Hz), alpha (8–12 Hz), beta (12–30 Hz), and gamma (30–70 Hz)—were analyzed for EEG power. The relative normalized power at each individual frequency is presented as a fraction of the sum of powers at all frequencies. The electrical activities were recorded after amplification (×1,200), bandpass filtering from 0.1 to 100 Hz and digitization at a 400 Hz sampling rate (AS 40) with a digital EEG system (Comet XL; Astro-Med, Inc., Warwick, RI, USA). EEG-video data were analyzed offline using PSG Twin (Astro-Med, Inc.) and Clampfit (Axon Instruments, Foster City, CA, USA).
Blood and tissue sampling
Anesthesia was induced by intraperitoneal injection of ketamine hydrochloride (100 mg/kg) and xylazine (10 mg/kg). Within 1 hour after maltreatment behavior tests, all mice were sacrificed and blood samples were obtained by heart puncture with heparin used as an anticoagulant. Samples were centrifuged for 3,000× g at 4℃ for 20 minutes, and plasma was stored at –70℃ until enzyme-linked immunosorbent assay (ELISA) tests. Brain tissues were also rapidly extracted within 1 hour after maltreatment behavior test and immediately frozen in liquid nitrogen.
Protein sample preparation
Total protein extracts were obtained from the whole brain. The frozen brains were weighed and suspended in 2 mL of lysis buffer, based on ice-cold phosphate-buffered saline (PBS) solution, pH 7.2 supplemented with a protease inhibitor cocktail, 1-mmol phenylmethylsulfonylfluoride, 0.5% Triton X-100, per gram of tissue. Tissue was disrupted using sonication and centrifuged at 13,000× g for 30 minutes at 4℃. The total protein concentrations in supernatants were quantified using bicinchoninic acid (BCA) colorimetric assay kit (Pierce BCA protein assay kit, cat.23225; Thermo Fisher Scientific, Waltham, MA, USA). The prepared protein samples were used for ELISA.
Enzyme-linked immunosorbent assay
Plasma corticosterone was measured using a competitive ELISA kit (ab108821; Abcam, Cambridge, UK) according to the manufacturer’s instructions. Briefly, plasma samples were diluted at 1:50 with assay buffer. A 25-μL sample and standard were added into the well, immediately followed by biotin corticosterone antigen and incubation at room temperature for 2 hours. After washing the well 5 times, streptavidin-peroxidase conjugate was added to each well and incubated for 30 minutes. After washing, substrate and stop solution were added. A VERSAmax microplate reader (Molecular Devices, San Jose, CA, USA) was used to measure signal intensity from the wells at 450 nm.
Oxytocin level in extracted blood plasma was measured using a competitive ELISA kit (ADI-900-153A; Enzo Life Sciences, New York, NY, USA). Briefly, plasma samples were diluted 1:4 with assay buffer and loaded in duplicate into wells with serially diluted oxytocin standards. After storage overnight at 4℃, the excess reagents were washed away, and the bound oxytocin phosphatase was incubated with substrate. After 1-hour incubation time, the enzyme reaction was stopped and the optical density read at 405 nm using A VERSAmax microplate reader.
For brain-derived neurotrophic factor (BDNF) assays, brain tissue was lysed, homogenized, and diluted to 5 μg/μL in order to quantify the concentration of BDNF using a commercial ELISA kit according to the manufacturer’s instructions (KA0331; Abnorva, Taipei, Taiwan). Briefly, brain homogenates and serial BDNF standards were loaded in duplicate into a microplate coated with rat-specific BDNF monoclonal antibodies and incubated at 37℃ for 90 minutes. A biotinylated detection polyclonal antibody from goat-specific BDNF was added at 37℃ for 60 minutes. After washing with PBS buffer, avidin-biotin-peroxidase complex was added. After unbound conjugates were washed away with PBS buffer, horseradish peroxidase (HRP) substrate 3,3'5,5'-tetramethylbenzidine (TMB) was added to visualize HRP enzyme reactions and incubated at 37℃ in the dark for 25 to 30 minutes. TMB was catalyzed by HRP to produce a blue product that changed to yellow after adding acidic stop solution. The absorbance at 450 nm was measured with a microplate reader, and BDNF concentration in brain tissue was assessed by comparing values to the BDNF standard curve.
Statistical analysis
All data are shown as mean ± standard error of mean. Analysis of variance (ANOVA) was used to conduct multiple comparisons of means. Student t-test was performed in order to determine significant differences between two means. The p-values of <0.05 were considered significant.
Results
Maltreatment behavior in the mouse
We developed a behavioral system that measures maltreatment or abusive behavior toward a conspecific to study antisocial behavior in the mouse. We used the perpetrator-victim paradigm and a modified two-compartment operant conditioning chamber with a single nose-poke hole (Figure 1A).
Normal male C57BL/6 perpetrator mice would put their noses into the hole several times but clearly stopped nose-poking behavior within 30 minutes after seeing the mouse in the victim compartment suffering from foot shocks (Figure 1B). However, normal male C57BL/6 mice exhibited high numbers of nose-pokes in the random and without foot-shock conditions (Figure 1B). There was a significant difference in total number of nose-pokes among the three foot-shock conditions for 120 minutes (with: n = 17, 5.35 ± 0.90; without: n = 16, 38.25 ± 6.75; random: n = 10, 28.40 ± 2.84) (F1,40 = 15.65, p < 0.01, one-way ANOVA) (Figure 1C). These results indicate that perpetrators can become conditioned to associate their actions with their conspecific’s distress.
To examine whether mPFC and ACC, which are considered to be important in affective and emotional responses for perceiving others’ pain or fear (i.e., empathic behavior), are involved in this behavior, we locally injected lidocaine (4%) through a cannula into mPFC or ACC. With foot shocks, perpetrators injected with lidocaine into the mPFC exhibited increased nose-poking compared with control mice that were similarly treated with saline (Figure 1D). Such increased levels were also observed in perpetrators injected with lidocaine into the ACC (Figure 1D). There was a significant difference in the total number of nose-pokes among the three groups for 120 minutes (saline: n = 16, 7.13 ± 1.52; mPFC-lidocaine: n = 11, 16.10 ± 2.62; ACC-lidocaine: n = 11, 19.73 ± 3.36) (F1,35 = 7.87, p < 0.01, one-way ANOVA) (Figure 1E). These results demonstrate that neural functions in mPFC and ACC are involved in maltreatment behavior.
Taken together, these results suggest that perpetrators may, at their own will, empathically stop nose-poking behaviors in order not to cause distress to conspecific victims.
Strain difference in maltreating behavior
We additionally tested BALB/c and 129S4/Sv mouse strains and found that BALB/c mice showed a similar number of nose-pokes (Figure 2A and B), but 129S4/Sv mice (n = 11) displayed a low number of nose-pokes compared with C57BL/6 (n = 17) and BALB/c mice (n = 11) (F1,36 = 3.78, p < 0.05, one-way ANOVA) (Figure 2D), suggesting variations in maltreatment behavior among different inbred mice strains.
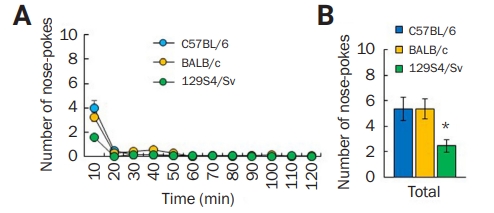
Maltreatment behavior of inbred mouse strains
(A, B) Number of nose-pokes by male C57BL/6 (n = 17), BALB/c (n = 11), and 129S4/Sv (n = 11) mouse strains every 10 minutes. (A) Average number of nose-pokes for a total of 120 minutes. (B) Among the strains, 129S4/Sv mice exhibited reduced numbers of nose-pokes compared to the others. *p < 0.05, one-way analysis of variance.
Molecular changes after maltreating behavior
Oxytocin, corticosterone, and BDNF levels are known to change with social stress and during prosocial or empathic behavior in human and animals. To determine whether those proteins are involved in maltreatment behavior, oxytocin, corticosterone, and BDNF levels in the perpetrators’ blood and brain were measured after maltreatment behavior. We found that perpetrator mice subjected to maltreatment behavior (maltreated group: n = 14, 139.88% ± 11.71%) had higher levels of oxytocin compared with the non-subjected mice (not maltreated group: n = 8, 100% ± 9.72%; p < 0.05, Student t-test) (Figure 3A). Corticosterone level also increased in the maltreated group (n = 15, 221.27% ± 18.22%) compared with the not maltreated group (n = 14, 100 %± 8.56 %; p < 0.01, Student t-test) (Figure 3B). However, the maltreated group (n = 15, 63.28% ± 1.94%) showed reduced BDNF level compared to the not maltreated group (n = 9, 100% ± 3.31%; p < 0.01, Student t-test) (Figure 3C). These results indicate the involvement of oxytocin, corticosterone, and BDNF signaling in maltreatment behavior.
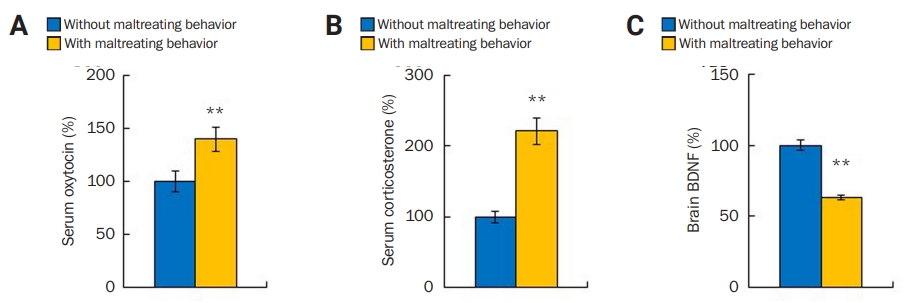
Alterations in oxytocin, corticosterone, and BDNF levels
(A-C) After maltreatment behavior, perpetrator mice in the condition with foot shocks had increased amounts of oxytocin (A) and corticosterone (B) but decreased BDNF level (C) compared to control mice in the condition without foot shocks. **p < 0.01, Student t-test.
BDNF, brain-derived neurotrophic factor.
Maltreatment behavior in the maternal separation/social defeat/social isolation model
Next, we used MS/SD/SI model mice as perpetrators in the maltreatment behavior task [53]. MS/SD/SI mice (n = 46) continued nose-poking behavior for the entire 120-minute experimental period and exhibited a higher number of nose-pokes compared to the control mice (n = 17) (F1,61 = 16.44, p < 0.01, two-way ANOVA) (Figure 4A). To compare nose-poking behavior according to social stress paradigm, the MD/SD or SI mice were also used in the maltreatment-behavior task. MS/SD (n = 11) and SI model mice (n = 15) showed similar numbers of nose-pokes to the control mice (Figure 4A). MS/SD/SI model mice (29.80 ± 5.43) exhibited much higher total number of nose-pokes among the models (control, 5.35 ± 0.90; MS/SD, 9.00 ± 2.53; SI, 8.33 ± 1.80) (F2,85 = 5.13, p < 0.01, one-way ANOVA) (Figure 4B). These results indicate that MS/SD/SI mice exhibit unique behavior in maltreatment-behavior tasks and suggest a cumulative effect of social stress on antisocial behavior in the mouse.
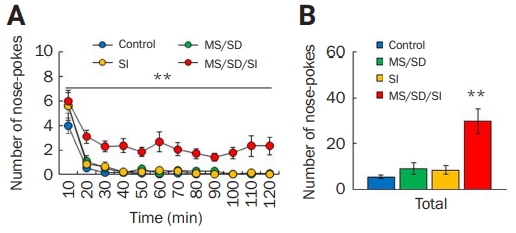
Maltreatment behavior in the MS/SD/SI model
(A, B) The number of nose-pokes by each MS/SD (n = 11), SI (n = 15), or MS/SD/SI mouse (n = 46) every 10 minutes (A) and for 120 minutes (B). Among the social stress models, only the MS/SD/SI model mice displayed high numbers of nose-pokes. **p < 0.01, two-way analysis of variance.
MS, maternal separation; SD, social defeat; SI, social isolation.
Differences in brain oscillations during maltreatment behavior
Many neuropsychiatric or personality disorders with deficits in empathy and social behavior, such as schizophrenia, autism, dementia, and psychopathy, show abnormal neural activities [38,39,55-62]. Thus, we investigated brain oscillations in MS/SD/SI model mice during maltreatment behavior. The control mice showed increased theta- (p < 0.01, Student t-test) and decreased alpha-rhythms (p < 0.05, Student t-test) after the nose-poking behavior compared to before the nose-poking (Figure 5A). However, the MS/SD/SI model mice exhibited a decrease in theta rhythm (p < 0.05, Student t-test) and an increase in alpha rhythm (p < 0.01, Student t-test) after the nose-poking behavior, opposite that of the control (Figure 5B). In addition, MS/SD/SI mice displayed decreased gamma-rhythm compared to control mice (p < 0.05, Student t-test) (Figure 5B).
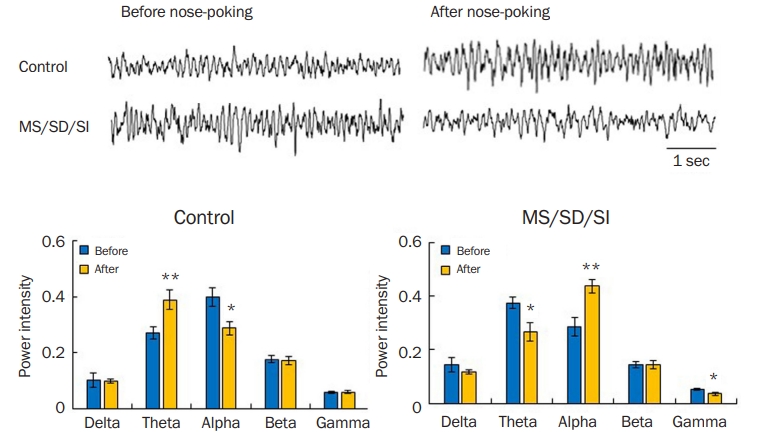
Brain oscillations during maltreatment behavior
The control mice (left, n = 11) showed increased theta- and decreased alpha-rhythms immediately after nose-poking behavior. However, the reverse pattern was observed in the MS/SD/SI model mice compared with the control (right, n = 13). Comparison between before and after power intensity; *p < 0.05, **p < 0.01, Student t-test.
MS, maternal separation; SD, social defeat; SI, social isolation.
Discussion
In this study, we developed an assay for studying antisocial maltreatment behavior in mice. We found that MS/SD/SI model mice generated by early-life chronic social stress paradigms (MS plus SD plus SI) are a suitable candidate mouse model of child abuse or maltreatment.
In the maltreatment-behavior task, maltreatment behaviors of perpetrators in the condition with foot shocks were clearly different from their behaviors in the condition without foot shocks or the condition with random foot shocks. However, there was no difference in the number of nose-pokes between the condition without foot shocks and the condition with random foot shocks. These imply that the reduced number of nose-pokes of the perpetrators in the condition with foot shocks is due to reduced locomotion or motor activity of perpetrators by observational fear.
In our pilot experiment, lower current intensities (0.2–0.6 mA) tended to elicit a high number of nose-pokes compared with 1-mA intensity (data not shown), implying that the behavioral response of the victim mouse affects nose-poking behavior of the perpetrator mouse. The perpetrator mouse becomes conditioned to, or learns, fear by observing the behavior of a conspecific victim and perceives the situation in which they may elicit noxious or aversive stimuli to the conspecific. In addition, inactivation of mPFC or ACC, which are considered to be engaged in affective and emotional responses for perceiving others’ pain or fear in animals and humans [39,63-69], worsened the maltreatment behavior of perpetrators. The difference only during the first 20 minutes of the session is probably due to the acute single injection of lidocaine. Taken together, these results suggest that mice can understand, or learn, the principle of the relationship between nose-poking behavior and foot shocks in the perpetrator-victim paradigm, and they may perceive the conspecific victim’s pain or fear in the opposite chamber, and thus they, by their own will, might stop nose-poking in order not to give an aversive stimulus, or damage, to a conspecific victim.
We observed differences between mouse strains in the maltreatment-behavior task. First, 129Sv mice exhibited a low number of nose-pokes compared with C57BL/6 and BALB/c mice. Previous animal studies have suggested strain differences in empathic behavior and aggressiveness [70-73]. In a previous study, we observed that MS/SD/SI mice showed increased offensive aggressiveness, reduced predator fear, and impairment in empathy-related behavior [53]. These behavioral traits may be involved in the antisocial maltreatment behavior of the MS/SD/SI mice. In this context, the maltreatment-behavior task may be a good behavioral task to measure antisocial behavior related to empathy deficit.
In conclusion, we developed a maltreatment-behavior task as an assay for antisocial behavior related to empathy deficit. The MS/SD/SI mouse model may be used to better understand personality disorders such as psychopathy showing antisocial behavior, impaired empathy, and excessive boldness. Cumulative social stress in early life may make mice susceptible to antisocial abusive behaviors. Further studies of maltreatment behavior following treatment with drugs acting on mental disorders would help to characterize the phenotypes of MS/SD/SI model mice as well as the maltreating assay in more detail.
Notes
Conflicts of Interest
Daejong Jeon has been an Associate Editor of encephalitis since October 2020. Kon Chu and Sang Kun Lee also have served on the editorial board of encephalitis since October 2020. They were not involved in the review process of this original article. No other potential conflict of interest relevant to this article was reported.
Author Contributions
Conceptualization, Funding acquisition, Resources, Project administration, Supervision: Jeon D, Lee SK, Chu K; Methodology, Investigation, Data curation: Jeon D, Choi J, Yang AR, Kim S; Formal analysis, Validation, Visualization: Jeon D, Choi J, Kim S; Writing-original draft: Jeon D; Writing-review & editing: Jeon D, Lee SK, Chu K.
Acknowledgements
We would like to thank Young-Sook Kim and Jung-Seok Yoo (Biomedical Research Institute, Seoul National University Hospital, Seoul, Korea) and their help with the ELISA and behavioral tests, respectively. This work was supported by a grant from Advanced Neural Technologies (0620182930).